厌氧氨氧化(anaerobic ammonia oxidation, ANAMMOX)技术是目前已知的最简单、最经济的生物脱氮技术, 自1995年被发现后就备受研究学者的关注.到目前为止, 人类已经发现了厌氧氨氧化菌的5个属, 其中只有1个属(Candidatus scalindua)是海洋性的, 且研究已证实Candidatus scalindua在海洋氮循环过程中发挥着关键性的作用:Ward等[5]研究发现海洋中30%~50%的N2 产出与厌氧氨氧化菌有关; Dalsgaard等研究发现由海洋厌氧氨氧化活动产生的N2 量约占全球N2 生成量的1/3~2/3;Thamdrup等用15N元素示踪法研究表明波罗的海沉积物中高达67%的N2 生成和海洋厌氧氨氧化作用相关. Schouten等发现海洋厌氧氨氧化菌能影响无机碳的固定, 从而对全球气候变化产生重要影响.因此对海洋厌氧氨氧化研究的不断深化不仅有利于海洋氮循环系统的完善, 而且能为全球气候的变化提供理论依据.
许多行业废水, 如水产养殖废水、海产品加工废水等, 盐度较高, 不利于淡水厌氧氨氧化菌的生长, 使高盐废水的脱氮处理更困难. Kartal等研究发现经海水驯化可培养出耐盐性的淡水厌氧氨氧化菌, 但驯化时间长, 且盐度一旦超过30 g・L-1, 就会强烈抑制淡水厌氧氨氧化菌的活性, 使脱氮效果降低.与淡水厌氧氨氧化菌相比, 海洋厌氧氨氧化菌本身生长的环境就是高盐度的海洋环境(30‰~35‰, 30~35 g・L-1), 不需要经过漫长的盐度驯化就能在30 g・L-1以上的盐度环境中发挥高效的厌氧氨氧化活性, 因此可用其直接处理高盐废水, 使高盐废水的脱氮处理更加简单易行.
虽然海洋厌氧氨氧化菌广泛存在于海洋环境中, 但分布却十分分散, 很难直接从海洋环境中得到高浓度的菌种, 这就对海洋厌氧氨氧化菌在废水处理方面的实际应用造成困难, 因此探究如何有效地富集培养海洋厌氧氨氧化菌就显得格外重要.目前国内对这方面研究主要还是体现在淡水厌氧氨氧化菌的富集培养上, 对海洋厌氧氨氧化菌的富集培养却鲜见报道.本试验以黄海胶州湾的底泥作为接种污泥, 进水为全海水, 在25℃条件下探究了海洋厌氧氨氧化菌的富集培养及其脱氮特性, 以期为海洋厌氧氨氧化在高盐废水处理方面的实际应用提供理论和技术指导.
1 材料与方法1.1 接种污泥
本试验使用的污泥是来自胶州湾的底泥, 黑色泥状, 带有臭味.污泥在进入反应器之前首先要进行淘洗, 去除里面的杂质.淘洗液的成分为0.01 g・L-1的KH2PO4、0.005 6 g・L-1的CaCl2与0.3 g・L-1的MgSO4.用淘洗液反复淘洗3次, 每次15 min, 将其静置30 min, 待污泥沉淀完全后倒掉上层的淘洗液, 然后将淘洗后的污泥倒入ASBR反应器.
1.2 试验用水
本试验用海水(5L)作为ASBR反应器的进水, 加入0.029 g・L-1的KH2PO4、0.136 g・L-1的CaCl2、1.2 g・L-1的KHCO3和0.3 g・L-1的MgSO4・7H2O, 微量元素Ⅰ和Ⅱ各1 mL・L-1.其中微量元素Ⅰ的成分及含量:EDTA为5 g・L-1, FeSO4・7H2O为5 g・L-1; 微量元素Ⅱ的成分及含量:EDTA为15 g・L-1, H3BO4为0.014 g・L-1, MnCl2・4H2O为0.99 g・L-1, CuSO4・5H2O为0.250 g・L-1, ZnSO4・7H2O为0.430 g・L-1, NiCl2・6H2O为0.190 g・L-1, CoCl2・6H2O为0.24 g・L-1, NaSeO4・10H2O为0.21 g・L-1.海水取自黄海(青岛麦岛周边), NH4+-N与NO2--N分别由NH4Cl与NaNO2按需提供, 将进水NH4Cl与NaNO2的初始浓度均设置为110 mg・L-1, 用1 mol・L-1的盐酸将进水pH控制在7.5±0.1范围, 进水用高纯氮气(纯度为99.99%)曝气15 min后导入ASBR反应器.
1.3 试验装置
试验用序批式厌氧活性污泥反应器(anaerobic sequencing batch reactor, ASBR)作为海洋厌氧氨氧化菌富集培养过程的反应器, 进水(3 min)、反应(10 h/24 h)、沉淀(45 min)、出水(2 min).反应器的结构如图 1所示:该反应器是由有机玻璃制成的圆柱体, 总体积为8 L, 有效体积为7 L.器壁的垂直方向上设有3个取样口, 取样口与排水口处的橡胶管均用夹子夹住, 防止空气中的O2进入反应器.曝完气的海水通过进水泵导入反应器, 出水通过出水管排出.反应器内设电动搅拌装置, 机械搅拌速率为80 r・min-1, 外部为水浴控温系统, 将温度控制在25℃左右.整个反应器的外围包裹一层黑布, 以防止光对海洋厌氧氨氧化菌产生抑制作用.

1.取样口及进出水口; 2.排泥口; 3.搅拌; 4.水浴循环进出水口; 5.水浴恒温控制器; 6.水封; 7.进水蠕动泵; 8.水浴循环蠕动泵; 9.储水箱
图 1 ASBR反应器装置示意
1.4 分析项目与方法
氨氮:纳氏试剂分光光度法; 亚硝酸盐氮:N-(1-萘基)-乙二胺光度法; 硝酸盐氮:麝香草酚分光光度法; pH/ORP:德国WTW pH/ORP/Oxi 340便携式pH计; 温度:水银温度计; 紫外/可见分光光度计:UV-5200.
SEM检测:在沉淀中加入2.5%的戊二醛5 mL, 置于4℃冰箱中固定4 h; 用0.1 mol・L-1, pH为8.0的磷酸缓冲溶液冲洗3次, 每次10 min; 分别用浓度为30%、50%、70%、90%的乙醇进行脱水, 每次15 min, 再用100%的乙醇脱水3次, 每次15 min; 然后加入100%乙醇与乙酸异戊酯的混合液(体积比为1:1) 及纯乙酸异戊酯各一次进行置换, 每次15 min; 对样品真空干燥后, 喷金, 通过扫描电镜(HITACHIS-4300) 观察污泥形态.
2 结果与讨论2.1 海洋厌氧氨氧化菌富集培养过程
根据反应器中海洋厌氧氨氧化菌的脱氮特性将整个富集培养过程分为4个阶段:菌体自溶阶段(1~15 d)、活性迟滞阶段(16~152 d)、活性提高阶段(153~183 d)与稳定运行阶段(184~192 d).
2.1.1 菌体自溶阶段(1~15 d)
反应器水力停留时间(HRT)设置为24 h, NH4+-N与NO2--N的起始浓度均为110 mg・L-1, 历经15 d, 完成菌体自溶期.如图 2所示:出水NH4+-N的浓度高于进水, 对NH4+-N的去除率为负值, 但随着反应器的运行, 进出水NH4+-N的浓度差逐渐减小至零; 对NO2--N的去除率明显下降, 仅经过8 d, 就由第1 d的84.44%降至24.25%;该阶段几乎没有NO3--N生成.出现该现象的原因是:刚进入反应器的菌体不适应新环境自溶释放出有机氮, 进而被分解成NH4+-N, 而厌氧条件下, 唯一消耗NH4+-N的海洋厌氧氨氧化菌数量极少, 活性极弱, 使NH4+-N的消耗量小于生成量; 在接种污泥中存在着分布广泛的异养反硝化菌, 它们以死亡的菌体作为有机碳源, 将NO2--N与NO3--N转化成N2, 使出水NO2--N与NO3--N的浓度下降.在8~15 d, 对NO2--N的去除速率较1~8 d平缓, 可能是菌体已适应新环境, 自溶作用减弱, 释放的有机碳减少, 使反硝化菌活性下降, 对NO2--N与NO3--N的转化速率下降.第15 d, 对NO2--N的去除率已降至10.78%, 此时出水NH4+-N的浓度为116.10mg・L-1, 低于进水, 对NH4+-N的去除率为0.6%, 开始出现正值, 这标志着菌体自溶阶段结束.
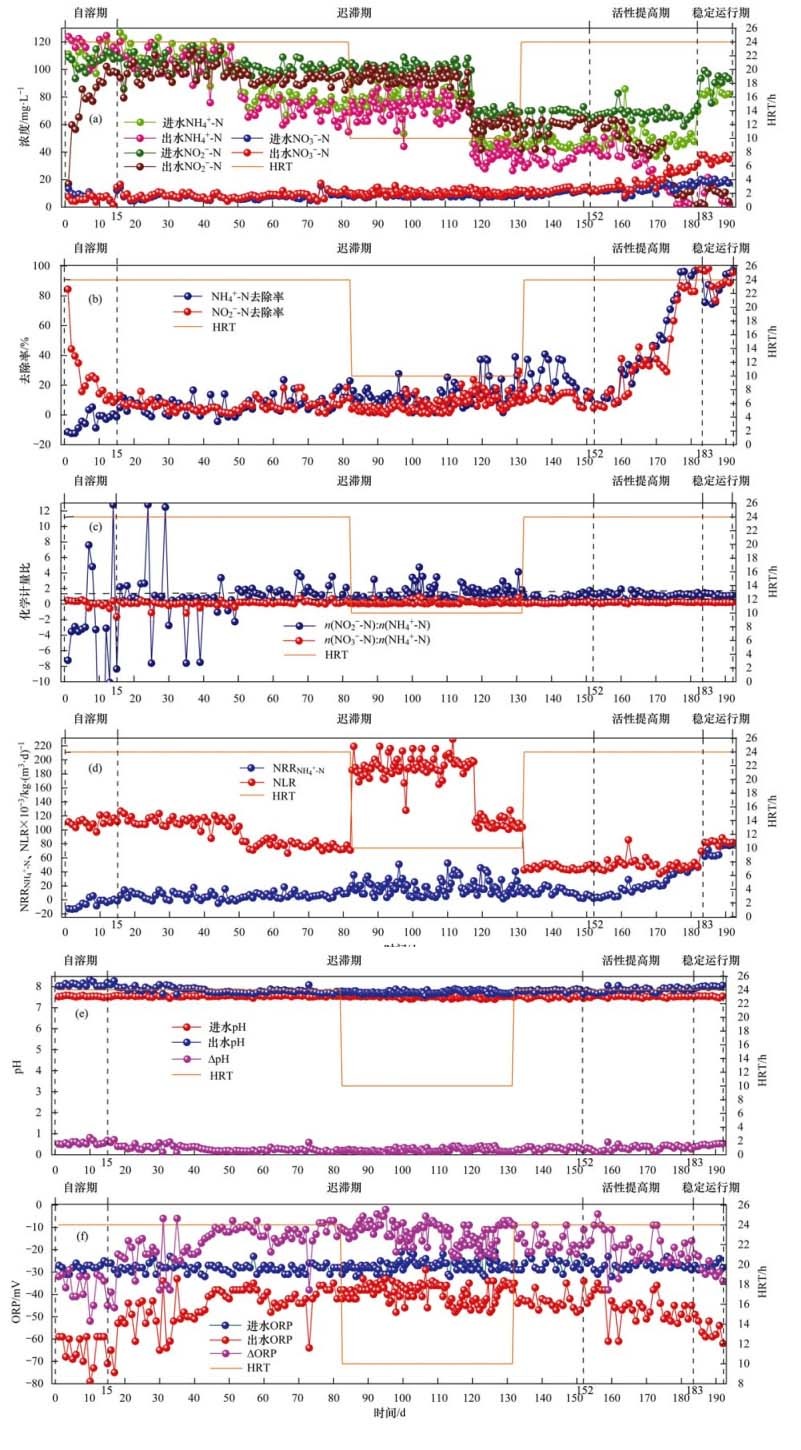
图 2 富集培养期间反应器主要指标变化
该阶段n(NO2--N):n(NH4+-N)为负值, 且波动很大, 只有少数几天出现正值情况; n(NO3--N):n(NH4+-N)很小, n(NH4+-N):n(NO2--N):n(NO3--N)未呈现一定的规律, 与理论比(1:1.32:0.26) 相差甚远, NRRNH4+-N为负值, 表明此阶段海洋厌氧氨氧化菌活性极低, 几乎为零, 异养反硝化菌占反应器主体.出水pH为8.03~8.34, ΔpH为0.48~0.81, 差值较大, 这是由于该阶段反硝化作用强烈, 产生大量碱度的缘故.进水ORP为-25~-31 mV, 出水ORP值为-59~-79 mV, 波动大.该阶段污泥仍为黑色泥状, 且污泥表现出来的特性与淡水厌氧氨氧化菌富集培养过程表现出来的特性一致.
2.1.2 活性迟滞阶段(16~152 d)
如图 2所示:16~50 d, NH4+-N的去除率不稳定, 波动很大, 甚至出现负值情况, 但大多数情况下为正值, 且去除率很低; 期间NH4+-N去除率最大值出现在第37 d, 为16.61%, 到第50 d, 去除率仅为1.35%;NO2--N的去除率总体呈下降趋势, 第16 d, 去除率为13.05%, 第47 d, 去除率降至1.63%;该阶段有少量NO3--N生成, 平均生成量为0.77 mg・L-1.这是由于NH4+-N的去除仅仅依赖海洋厌氧氨氧化菌的作用, 但该菌数量少, 活性低, 且该阶段依然存在菌体自溶现象, 导致NH4+-N的去除率低且不稳定; 对NO2--N的去除依赖海洋厌氧氨氧化菌与异养反硝化菌的共同作用, 该阶段异养反硝化菌仍占优势, 因此对NO2--N的去除贡献最大的反应依然是反硝化反应, NO2--N去除率下降标志着反硝化菌活性下降. n(NH4+-N):n(NO2--N):n(NO3--N)波动大, 未呈现一定的规律, 且n(NO2--N):n(NH4+-N)与n(NO3--N):n(NH4+-N)均小于理论值, 这可能是异养反硝化菌将NO3--N转化成NO2--N, 减少了NO2--N的净消耗量与NO3--N的生成量, 造成NO2--N的累积. NRRNH4+-N平均值为0.005 4 kg・(m3・d)-1, 很低且波动大, 有时甚至出现负值, 表明海洋厌氧氨氧化菌活性及其微弱.出水pH为7.7~8.0, 较自溶期有所下降.到第50 d, NH4+-N和NO2--N的去除率分别降至1.35%与2.62%, 可能是进水基质浓度(NH4+-N与NO2--N均为110 mg・L-1)过高对海洋厌氧氨氧化菌的活性产生抑制所致.该过程初步表明海洋厌氧氨氧化菌对高浓度的进水基质较敏感, 基质浓度过高不利于海洋厌氧氨氧化菌的富集培养.
考虑到高浓度基质对菌的抑制作用, 从第51 d开始, 按海洋厌氧氨氧化反应方程式中NH4+-N与NO2--N的消耗比(1:1.32) 配水, 并将进水NH4+-N与NO2--N的浓度分别降至80mg・L-1与105.6mg・L-1.如图 2所示:反应刚开始, NH4+-N与NO2--N的去除率均略微升高(3%左右), 但并未出现持续升高现象, 而是有明显波动, 但总体呈小幅度上升趋势, 第82 d, NH4+-N与NO2--N的去除率分别为22.88%与11.15%;且该阶段有明显的NO3--N生成, 平均生成量为1.51mg・L-1. n(NO2--N):n(NH4+-N)基本高于1.32, 而n(NO3--N):n(NH4+-N)基本低于0.26, 且波动均较大, 表明反硝化菌与海洋厌氧氨氧化菌对NO2--N的总消耗量大于反硝化作用NO2--N的生成量. NRRNH4+-N波动变小, 且有升高的趋势, 但基本都在0.01 kg・(m3・d)-1以下.出水pH为7.7~7.9, ORP值为-38~-46 mV, 均比高基质浓度的波动范围小, 说明降低进水基质浓度有利于海洋厌氧氨氧化菌的富集.
淡水厌氧氨氧化菌活性提高的方法有两个, 一是提高反应基质的浓度; 二是缩短水力停留时间(HRT).因为海洋厌氧氨氧化菌在迟滞期初期就初步显示了对较高浓度基质敏感性的特点, 所以本试验考虑用第二种方法提高菌的活性.在第82 d, 将HRT由24 h缩短到10 h, 其余条件不变.如图 2所示:NH4+-N去除率只是略微上升, 但随后并未出现预期的持续上升现象, 而是出现明显波动, 且呈下降趋势, 到第107 d, 去除率降至7.66%, 猜想可能是HRT缩短使反应器中氮容积负荷(NLR)过高, 对海洋厌氧氨氧化菌产生较大冲击, 使脱氮效果变差.随后NH4+-N去除率略微上升, 到117 d, 升至13.73%;第82~111 d, 对NO2--N的去除率基本在10%以下, NO3--N生成量在1mg・L-1以下; 随后NO2--N的去除量与NO3--N生成量总体呈上升趋势, 到117 d, NO2--N去除率升至23.61%, NO3--N生成量为4.64 mg・L-1.该阶段NRRNH4+-N波动大[0.003 4~0.051 kg・(m3・d)-1], 但总体呈先升高后降低, 随后又升高的趋势, 第117 d, NRRNH4+-N为0.027 kg・(m3・d)-1.该阶段初步表明海洋厌氧氨氧化菌抗氮容积负荷的冲击能力差, 对HRT变化较敏感.
第118 d, 保持HRT不变, 将进水NH4+-N与NO2--N的浓度分别降至50 mg・L-1与66 mg・L-1, 通过降低基质浓度来缓解高NLR对海洋厌氧氨氧化菌的冲击影响.如图 2所示:反应前期NH4+-N的去除率逐渐升高, 到122 d, 升至36.79%, 随后去除率逐渐下降, 仅经过4 d, 就降至1.6%, 最后又略微升高, 第131 d, 去除率为16.52%;NO2--N去除量和NO3--N的生成量与NH4+-N的变化一致. NRRNH4+-N仍波动较大[0.001 7~0.046 kg・(m3・d)-1], 说明此时海洋厌氧氨氧化菌活性仍不稳定, 依然处于活性迟滞阶段.该现象表明短HRT造成的高NLR会对海洋厌氧氨氧化菌的活性产生抑制, 且不能通过降低基质浓度的方法消除.因此, 第132 d保持进水基质浓度不变, 将HRT重新延长至24 h, 如图 2所示:第132~143 d, NH4+-N与NO2--N的去除率总体升高, 143 d, 分别升至36.79%与14.57%, NRRNH4+-N由0.009 2 kg・(m3・d)-1升至0.016 kg・(m3・d)-1, 随后NH4+-N与NO2--N的去除率均下降, 到152 d, 分别降至4.8%与4.64%, NRRNH4+-N由0.011 kg・(m3・d)-1降至0.002 8 kg・(m3・d)-1, 从153 d开始, NRRNH4+-N持续上升, 这标志着菌体迟滞期结束.
海洋厌氧氨氧化菌富集培养过程中活性迟滞时间为137 d, 占整个富集培养时间的2/3以上. Yu等用厌氧污泥富集培养淡水厌氧氨氧化菌时, 活性迟滞时间为30 d; Tang等用UASB反应器富集培养淡水厌氧氨氧化菌的迟滞期为29 d; Wang等用好氧硝化污泥富集培养淡水厌氧氨氧化菌时, 迟滞期仅有20 d.由此可见, 海洋厌氧氨氧化菌富集培养过程中活性迟滞时间要远长于淡水厌氧氨氧化菌, 这使海洋厌氧氨氧化菌的富集培养更难.
2.1.3 活性提高阶段(153~183 d)
从153 d开始, NH4+-N与NO2--N的去除率均持续升高, 第155 d, 去除率分别为11.76%与5.9%, 第164 d, 去除率分别为30.18%与32.93%, 第174 d, 去除率分别为70.9%与50.91%, 第183 d, 出水NH4+-N的浓度仅为0.37 mg・L-1, NO2--N的浓度仅为0.323 mg・L-1, 去除率分别升至99.47%与99.54%, 几乎达到100%. NO3--N的生成量由0.668 8 mg・L-1升至14.11 mg・L-1; NRRNH4+-N由0.003 5 kg・(m3・d)-1升至0.069 kg・(m3・d)-1, 海洋厌氧氨氧化菌活性明显提高.该阶段n(NH4+-N):n(NO2--N):n(NO3--N)为1:1.15:0.23, 接近理论比(1:1.32:0.26) , 这标志着海洋厌氧氨氧化菌已完全取代异养反硝化菌, 成为反应器的主体菌.出水pH为7.7~7.9, ORP为-37~-43 mV.
本试验仅用30 d, NH4+-N与NO2--N的去除率就从7.19%与6.87%分别升至99.47%与99.54%, 活性提高期低于整个富集培养时间的1/6.赖玮毅等历经60 d才使淡水厌氧氨氧化菌活性显著提高, Xing等用污水处理厂的活性污泥富集培养淡水菌时, 活性提高期为69 d.因此前者在活性提高阶段比后者历时短.
2.1.4 稳定运行阶段(184~192 d)
第184 d, 将进水NH4+-N与NO2--N的浓度分别升至80 mg・L-1与105.6 mg・L-1, 如图 2所示:NH4+-N去除率稳定上升, 从起初的75.52%提升到96.98%, 而NO2--N的去除率均稳定在85%以上, 最高去除率能达到98.4%. n(NH4+-N):n(NO2--N):n(NO3--N)为1:(1.2±0.2):(0.22±0.06), 接近理论比(1:1.32:0.26) . NRRNH4+-N由0.062 kg・(m3・d)-1升至0.080 kg・(m3・d)-1, 出水pH为7.9~8.06, ORP为-49~-62 mV.
本试验用黄海胶州湾的底泥, 经192d成功富集培养了海洋厌氧氨氧化菌, 在稳定运行期, NH4+-N与NO2--N的去除率分别为96.98%与95.66%. Gong等在富集培养淡水厌氧氨氧化菌时, 经120 d对NH4+-N与总氮的去除率就分别达到88.7%与83.77%;Chen等在UASB反应器中仅用40 d, 就实现了淡水厌氧氨氧化菌活性的显著提高, 此时NRR为0.55 kg・(m3・d)-1; Wang等经61 d实现了淡水厌氧氨氧化菌的富集, 对NH4+-N与NO2--N去除率均能达到91%以上.由此可见, 海洋厌氧氨氧化菌的富集培养时间要远长于淡水厌氧氨氧化菌, 但稳定运行时, 前者的脱氮效果更好.原因可能是海洋水体成分复杂, 含有众多元素及少量有机质, 其中含有的某种成分对海洋厌氧氨氧化菌的脱氮产生促进作用, 具体原因还有待探究.
2.2 不同阶段周期内反应器脱氮特性
海洋厌氧氨氧化菌的富集培养过程具有不同的阶段特征, 为了比较4个阶段沿程NH4+-N、NO2--N、NO3--N、pH与ORP的变化情况, 在第15、152、183与192 d, 每隔1 h测一次水样, 看反应器内各个指标的沿程变化情况.如图 3所示:在菌体自溶期, 反应器中NH4+-N的浓度沿程波动较大, 且未呈现一定的规律; NO2--N的浓度逐渐降低, 由109.31 mg・L-1降至97.53 mg・L-1, 变化较平缓, 去除量仅为11.78 mg・L-1; NO3--N生成量仅为0.23 mg・L-1, 此现象与菌体自溶作用有关; 迟滞期NH4+-N、NO2--N、NO3--N、pH与ORP沿程变化极平缓, 几乎无变化, 这是由于海洋厌氧氨氧化菌的活性延迟所致; 在活性提高期与稳定运行期, NH4+-N与NO2--N的沿程变化可分为3个阶段: ① 前5 h, 浓度下降较平缓, 可能是废水的进入改变了反应器内的环境, 海洋厌氧氨氧化菌对新环境不适应使其活性发生延迟所致, 本试验得出的菌活性延迟时间为5 h.于德爽等在研究淡水厌氧氨氧化菌富集培养时发现:由进出水导致的菌活性延迟时间仅为0.8 h, 王晓霞等在用2:1的好养短程硝化污泥与厌氧氨氧化污泥富集淡水厌氧氨氧化菌时, 发现该时间为2 h, 均小于5 h.由此可见, 海洋厌氧氨氧化菌抗水力冲击能力更弱, 对新环境的适应能力远小于淡水厌氧氨氧化菌. ② 5~19 h, 浓度下降迅速, 充分显示出海洋厌氧氨氧化活性. ③ 19~24 h, 浓度下降平缓, 可能是反应器中NH4+-N与NO2--N的浓度较低, 低浓度基质满足不了海洋厌氧氨氧化菌的生长需求, 使其活性降低, 对NH4+-N与NO2--N的去除速率下降.

图 3 周期内不同阶段各指标的变化情况
4个阶段pH值均沿程不断上升, 开始时, pH值分别为7.48、7.44、7.56、7.54, 周期结束时, pH值分别为8.02、7.55、7.83、8.06, ΔpH分别为0.54、0.11、0.27、0.52.由此可见:菌体自溶期与稳定运行期pH上升速率明显快于活性迟滞期与活性提高期, 可能是这两个阶段的主体菌分别是具有高活性的反硝化菌与海洋厌氧氨氧化菌, 它们分别发生强烈的反硝化作用与厌氧氨氧化作用, 产生的碱度最多, pH上升速率最快; 在迟滞期, 反硝化菌活性低, 且厌氧氨氧化菌活性延迟, 因此迟滞期的pH值上升速率最慢. ORP值的变化情况与pH类似, 4个阶段ORP值均沿程不断下降, 由开始的-25、-23、-29、-28 mV分别降至-58、-29、-45、-62 mV, ΔORP分别为-33、-6、-16、-34 mV; 菌体自溶期与稳定运行期ORP下降速率明显快于活性迟滞期与活性提高期, 其原理与pH值变化原理相同.
2.3 富集培养过程中污泥性状的变化
污泥进入反应器之前为黑色泥状[图 4(a)], 带有臭味, 经淘洗后接种到ASBR反应器, 运行51 d, 污泥由黑色变为黑褐色, 仍为泥状, 未形成颗粒, 污泥沉降性能差; 51~83 d, 污泥由黑褐色泥状变为褐色絮状[图 4(b)], 但污泥沉降性能依然很差; 第132 d, 反应器中开始出现隐隐约约暗红色絮状污泥; 132~173 d, 暗红色絮状污泥逐渐变成浅红色小颗粒状[图 4(c)], 污泥沉降性能好; 在稳定运行阶段, 污泥转化为砖红色大颗粒状[图 4(d)], 将此阶段的污泥用10倍显微镜观察, 如图 5(a)所示:该污泥为砖红色颗粒状的圆球体, 内部有少量黑色物质, 结构紧密, 边缘较光滑, 与文献报道的成熟的厌氧氨氧化菌的形态类似.经扫描电镜观察, 该污泥是由大量的球状菌经胞外多聚物黏聚而成菌团.因此, 同淡水厌氧氨氧化菌富集培养类似, 也可以用污泥性状的变化来反映海洋厌氧氨氧化菌的富集情况.具体参见污水宝商城资料或http://www.dowater.com更多相关技术文档。
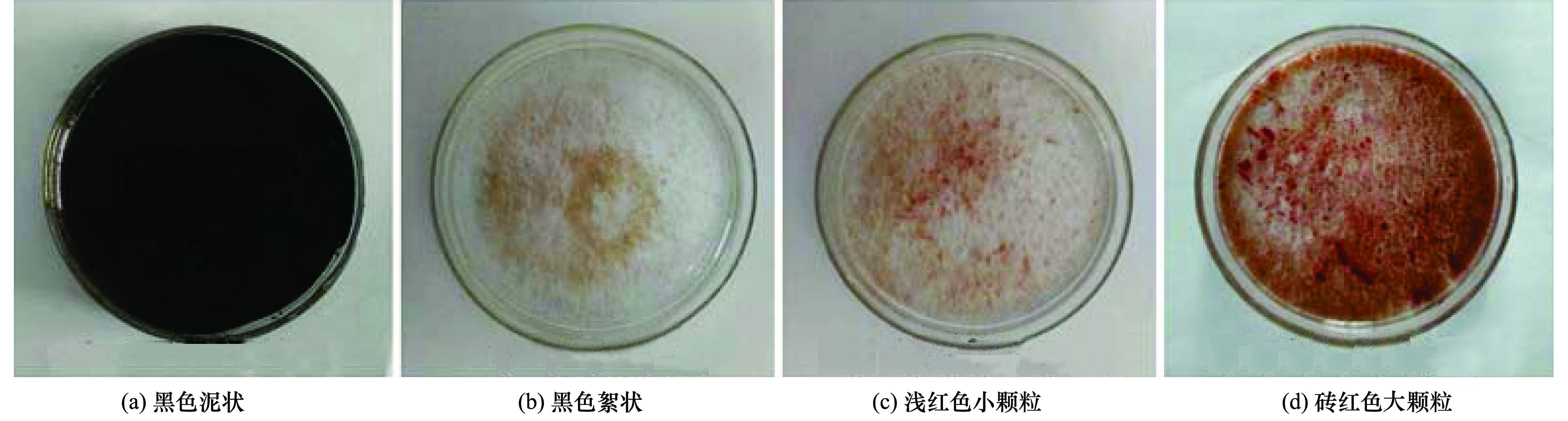
图 4 富集培养过程中污泥性状的变化情况
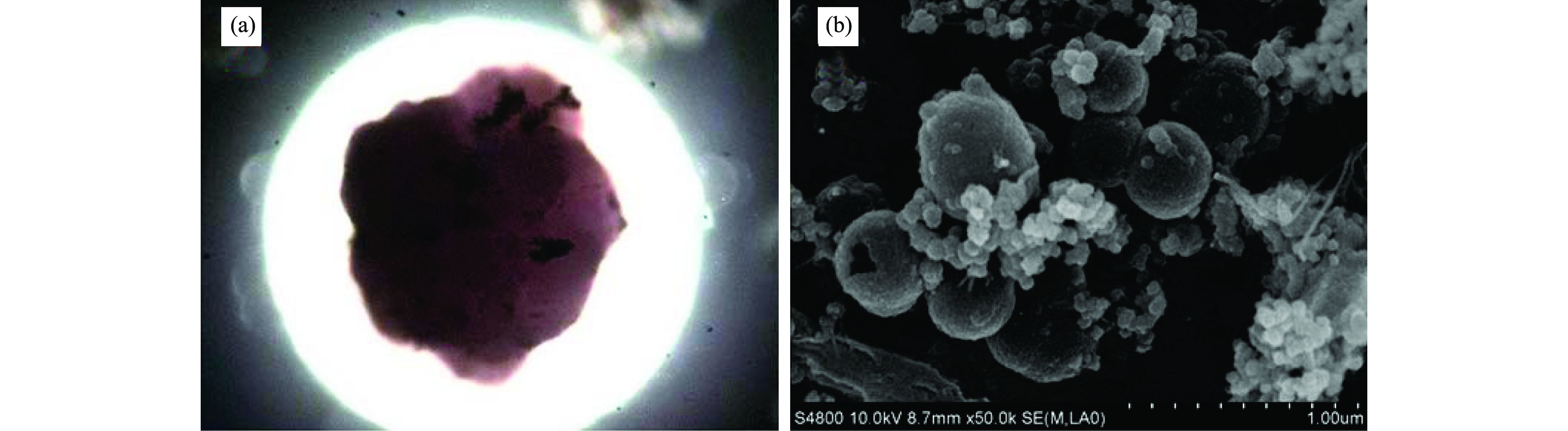
图 5 颗粒污泥的影像照片
3 结论
(1) 用黄海胶州湾的底泥, 经192 d成功富集培养了海洋厌氧氨氧化菌.富集过程分为4个阶段:菌体自溶期(1~15 d)、活性迟滞期(16~152 d)、活性提高期(153~183 d)与稳定运行期(184~192 d).富集成功后, NH4+-N与NO2--N的去除率分别为96.98%与95.66%.
(2) 与淡水厌氧氨氧化菌富集培养相比, 海洋厌氧氨氧化菌的迟滞时间(137 d)较长, 占整个富集时间的2/3以上, 活性提高期(30 d)较短, 小于富集时间的1/6;且海洋厌氧氨氧化菌对基质浓度与HRT变化更敏感, 由进出水导致的菌活性延迟时间为5 h, 远长于淡水厌氧氨氧化菌, 因此海洋厌氧氨氧化菌对新环境的适应能力更弱, 更难富集培养.
(3) 反应器运行过程中, 污泥逐渐由黑色泥状变为褐色絮状, 随后变为暗红色小颗粒状, 在稳定运行阶段, 污泥为砖红色颗粒状, 扫描电镜观察该砖红色颗粒为表面光滑, 排列紧密、有类似火山口形状的球状菌相互黏聚而成的菌团.

