摘要:硝化反应是由一类自养好氧微生物完成的,它包括两个步骤:第一步称为亚硝化过程,是由亚硝酸菌将氨氮转化为亚硝酸盐,亚硝酸菌中有亚硝酸单胞菌属、亚硝酸螺杆菌属和硝化球菌属;第二步称为硝化过程,由硝酸菌(包括硝酸杆菌属、螺菌属和球菌属)将亚硝酸盐进一步氧化为硝酸盐。
垃圾填埋场的渗滤液属高浓度氨氮废水,后期渗滤液的氨氮浓度达2000mg/L以上,如利用生物法脱氮,反硝化需7500mg/L以上的碳源,而渗滤液本身所能提供的碳源明显不足,外加碳源则会增加处理成本。因此,研究高效脱氮工艺具有重要意义。近些年来在生物脱氮理论方面有了许多进展,亚硝酸盐硝化反硝化受到重视,发现了厌氧氨氧化和好氧反硝化微生物的生物化学作用,从而为高浓度氨氮废水的高效生物脱氮提供了可能的途径。
1 传统生物脱氮原理
硝化反应是由一类自养好氧微生物完成的,它包括两个步骤:第一步称为亚硝化过程,是由亚硝酸菌将氨氮转化为亚硝酸盐,亚硝酸菌中有亚硝酸单胞菌属、亚硝酸螺杆菌属和硝化球菌属;第二步称为硝化过程,由硝酸菌(包括硝酸杆菌属、螺菌属和球菌属)将亚硝酸盐进一步氧化为硝酸盐。亚硝酸菌和硝酸菌统称为硝化菌,都利用无机碳化合物如CO32-、HCO3-和CO2作为碳源,从NH3、NH4+或NO2-的氧化反应中获取能量。
亚硝酸菌和硝酸菌的特性大致相似,但前者的世代期较短,生长率较快,因此较能适应冲击负荷和不利的环境条件;当硝酸菌受到抑制时,有可能出现NO-2积累的情况。
反硝化反应是由一群异养型微生物完成的,它的主要作用是将硝酸盐或亚硝酸盐还原成气态氮或N2O,反应在无分子态氧的条件下进行。反硝化细菌在自然界很普遍,多数是兼性的,在溶解氧浓度极低的环境中可利用硝酸盐中的氧作为电子受体,有机物则作为碳源及电子供体提供能量并被氧化稳定。
由于从反硝化获得的能量低于氧气还原所获取的能量,所以反硝化被认为仅在缺氧条件下发生。
从NH4+至NO2-的转化,经历了3个步骤、6个电子的转移,可见亚硝酸菌的酶系统十分复杂,而硝化过程则相对简单些,只经历了一步反应、2个电子的变化。因此也有人认为,亚硝酸菌往往比硝酸菌更易受到抑制。
反硝化反应一般以有机物为碳源和电子供体。当环境中缺乏有机物时,无机物如氢、Na2S等也可作为反硝化反应的电子供体,微生物还可以消耗自身的原生质进行所谓的内源反硝化。
C5H7O2N+4NO3-→5CO2+NH3+2N2↑+4OH-
可见内源反硝化的结果是细胞物质的减少,并会有NH3的生成,因此废水处理中均不希望此种反应占主导地位,而应提供必要的碳源。
硝化和反硝化反应的进行是受到一定制约的,一方面,自养硝化菌在大量有机物存在的条件下,对氧气和营养物的竞争不如好氧异养菌,从而导致异养菌占优势;另一方面,反硝化需要提供适当的电子供体,通常为有机物。上述硝化菌和反硝化菌的不同要求导致了生物脱氮反应器的不同组合,如硝化与反硝化由同一污泥完成的单一污泥工艺和由不同污泥完成的双污泥工艺。前者通过交替的好氧区与厌氧区来实现,后者则通过使用分离的硝化和反硝化反应器来完成。如果硝化在后,需要将硝化废水进行回流;如果硝化在前,需要外加电子供体,这就是传统脱氮工艺存在的问题和困难所在。
这种两难处境在氨氮浓度低的城市污水处理中表现得还不很明显,在高浓度氨氮废水生物脱氮处理中则表现得很突出。近些年来,人们试图从各个方面突破生物脱氮的困境,如开发亚硝酸硝化/反硝化脱氮工艺;与传统生物脱氮理论相反的一些生物过程被发现,例如发现了氨与亚硝酸盐/硝酸盐在缺氧条件下被同时转化为氮气的生物化学过程,这一过程被称为厌氧氨氧化(Anammox);好氧反硝化和异养硝化作用也被发现,好氧反硝化往往与异养硝化同时发生;在有氧条件下能够反硝化的细菌也被分离出来,其中有异养菌(Thiosphaerapantotropha和Alcaligenessp)及自养硝化菌。
2 亚硝酸硝化/反硝化工艺
在硝化反应中,一般认为硝酸盐是反应的主产物,而从氨向亚硝酸盐的转化一般认为是硝化过程(Nitrification)的速度控制步骤,但是出现亚硝酸盐积累的报道也很多。
人们认为,出现亚硝酸积累是有害的。为了减少亚硝酸的积累,许多研究人员进行了控制其积累的工艺条件的研究工作,并得到了有关自由氨可抑制亚硝酸积累的结论,其结果也得到了证实并被广泛接受。随后,开始把注意力放在通过亚硝酸硝化―反硝化缩短脱氮过程上,这种工艺的潜在优势在于:①节省25%的硝化曝气量。②节省40%的反硝化碳源。③节省50%的反硝化反应器容积。
这些对于高浓度氨氮废水的脱氮处理具有非常大的经济效益,特别是对于诸如垃圾渗滤液等碳源不足的废水更是如此。
在硝化系统中,与亚硝酸积累有关的因素包括:①自由氨的存在,②较高的pH值,③溶解氧浓度低,④温度的变化,⑤氨氮负荷高,⑥污泥龄长,⑦硝酸盐的还原。大多数研究人员认为自由氨浓度高(高pH值条件下)和溶解氧浓度低是亚硝酸盐积累的主要原因,指出亚硝酸积累的内在原因在于自由羟氨(NH2OH)的积累。根据对前人试验结果的分析,表明自由羟氨不应是亚硝酸积累的最终原因,自由羟氨积累主要受溶解氧、pH的控制。有高浓度氨氮废水需要处理的单位,也可以到污水宝项目服务平台咨询具备类似污水处理经验的企业。
然而,实现亚硝酸反硝化的成功报道并不多见。Jetten等人利用硝酸菌和亚硝酸菌在较高温度下生长速度的显著差异,通过控制温度和污泥停留时间,将在高温下生长速度较慢的硝酸菌从反应器中冲洗出去,使亚硝酸菌在反应器中占优势,从而将氨氧化控制在亚硝化阶段,这种工艺叫作SHARON工艺(SingleReactorforHighActivityAmmoniaRemovalOverNitrite)。但该工艺须在30~40℃的温度下进行,只对温度较高的污水如厌氧消化排水的脱氮处理有实际意义。对于垃圾渗滤液等废水,必须从控制溶解氧及pH值来实现稳定的亚硝酸反硝化脱氮。
3 同时硝化/反硝化(SND)
当好氧环境与缺氧环境在一个反应器中同时存在,硝化和反硝化在同一反应器中同时进行时则称为同时硝化/反硝化。同时硝化/反硝化不仅可以发生在生物膜反应器中,如流化床、曝气生物滤池、生物转盘;也可以发生在活性污泥系统中,如曝气池、氧化沟。
同时硝化/反硝化的活性污泥系统为今后简化生物脱氮技术并降低投资提供了可能性。但目前对SND现象的机理还没有一致的解释,一般认为三个主要机理是:①混合形态。由于充氧装置的充氧不均和反应器的构造原因,造成生物反应器形态不均,在反应器内形成缺氧/厌氧段。此种情况称为生物反应的大环境,即宏观环境。②菌胶团或生物膜。缺氧/厌氧段可在活性污泥菌胶团或生物膜内部形成,即微观环境。③生物化学作用。在过去几年中,许多新的氮生物化学菌族被鉴定出来,其中包括部分菌种以组团形式对SND起作用,包括起反硝化作用的自养硝化菌及起硝化作用的异养菌。
在生产规模的生物反应器中,完全均匀的混合状态并不存在。菌胶团内部的溶解氧梯度目前也已被广泛认同,使实现SND的缺氧/厌氧环境可在菌胶团内部形成。由于生物化学作用而产生的SND更具实质意义,它能使异养硝化和好氧反硝化同时进行,从而实现低碳源条件下的高效脱氮。
4 好氧反硝化
最初,反硝化被认为是一个严格的厌氧过程,因为反硝化菌作为兼性菌优先使用溶解氧呼吸,甚至在浓度低达0.1mg/L时也是如此,这样就阻止了使用硝酸盐和亚硝酸盐作为最终电子受体,不过这种限制只是对专性厌氧反硝化菌起作用。20世纪80年代后期以来,在生物脱氮生物学方面有了很大进展。人们曾多次观察到在没有明显缺氧段的活性污泥法中存在脱氮现象,发现了好氧反硝化菌:Pseudomonasspp,Alcaligenesfaecalis,ThiosphaeraPantotropha,这些好氧反硝化菌同时也是异养硝化菌,而传统上的硝化菌是化学自养型的。这样,这类细菌就可将氨在好氧条件下直接转化成气态产物。同时可以查看中国污水处理工程网更多技术文档。
生物学研究表明,在好氧和缺氧条件下Nitrosmonasspp能够通过硝酸盐的生物还原形成氧化氮和氧化亚氮。有人认为,在好氧条件下氧化氮和氧化亚氮产生速率依赖于亚硝酸盐浓度,而大多数人则认为这一速率与溶解氧浓度成反比。众多研究表明,Nitrosmonasspp的反硝化活动在低溶解氧条件下是明显的,但对Nitrobacterspp的反硝化能力研究得比较少。有人认为在好氧条件下,Nitrobacter菌株不能进行反硝化,某些菌株可以在无氧的丙酮酸、氨和硝酸盐的培养物中生长,丙酮酸和硝酸盐被消耗,在低溶解氧条件下生产的氧化氮可能参与到NADH的形成。
反硝化的初始基质可能是亚硝酸盐或硝酸盐,研究比较电子转移平衡可以确定初始基质是硝酸盐还是亚硝酸盐。氨氧化为亚硝酸盐产生2个电子,亚硝酸盐氧化为硝酸盐也产生2个电子,完全的亚硝酸盐还原需要3个电子,而完全的硝酸盐还原需要5个电子。因此,当亚硝酸盐被完全还原时,氧化氨产生氮气的最大可能因数为0.67N-molN2/N-molNH3,而对于硝酸盐这一因数为0.4,所以亚硝酸盐为反硝化的初始基质。
Muller等证明了好氧反硝化是与硝化相伴发生的。假设a是流向最终细胞色素C氧化酶的电子占氨氧化净放出电子的比例,则(1-a)就是用于完全亚硝酸盐还原的电子比例。氨氧化菌完成氧化还原反应为:
NH3+O2→NO2-+3H++2e-
a(0.5O2+2e-+2H+→H2O)
(1-a)(0.67NO2-+2.67H++2e-→0.33N2+1.33H2O)
剩下的(0.33+0.67a)亚硝酸盐(mol)/被氧化的氨(mol)根据总反应得:
NH3+(1.17+0.83a)O2→(0.33+0.67a)・
NO3-+(0.33-0.33a)N2+(0.33+0.67a)・
H++(1.33-0.33a)H2O
若以qN2和qO2分别表示氮气产生和氧气消耗的速率,则
qN2+qO2=(0.33-0.33a)/(1.17+0.83a)
或 a=(qO2-3.5qN2)/(2.5qN2+qO2)
显然,a的数值介于0~1。如果氨氧化菌的呼吸消耗硝酸盐,在0.3kPa溶解氧压力条件下流向最终细胞色素C氧化酶的电子流将是负值。所以,氨氧化仅可以供给呼吸的亚硝酸盐电子。
Muller等人得到的在不同溶解氧压力下好氧反硝化特征参数如表1所示。
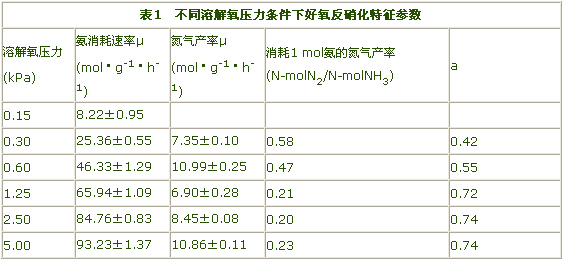 |
从表中可见,好氧反硝化速率与氨消耗速率基本处于同一数量级,这使好氧反硝化更具实际的工程意义,这将在节省能源消耗的情况下,使污水脱氮处理的效率大大提高。好氧反硝化在以往的研究与生产中之所以没有得到证实,主要是由于如下原因:其一,空气中含有高浓度氮气,要通过产气监测证实好氧反硝化的存在需要使用氮同位素、无氮载气、密闭反应器和质谱仪;其二,好氧反硝化菌浓度要足够高才能检测到氮气的产生,由于Muller研究中的氨氧化菌为0.3g/L、约占活性污泥的10%,而在一般活性污泥中,好氧反硝化菌Nitrosomonasspp仅有10mg/L,结果是产气量太低而检测不到;其三,过去研究的细菌常常是不能产生氮气的NitrosomonaseuropaeaATTC19718。
5 厌氧氨氧化(Anammox)
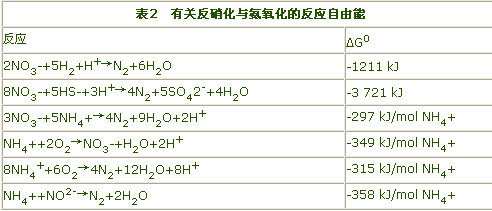
在理论上,氨也可以作为反硝化的无机物电子供体,其时的反应自由能几乎与好氧硝化的一样有利(参见表2)。早在1997年,Broda就发表了一篇题为“自然界中遗失的两种微生物”之论文,指出在自然界中可能存在一些微生物(chemolithotrophicbacteria)能够以硝酸盐、二氧化碳和氧气为氧化剂将氨氧化为氮气。根据这个预言,目前已经发现Nitrosomonaseutropha能够进行这种生物化学反应。
 |
Mulder在实验室规模的反硝化流化床反应器中,发现了氨和硝酸盐的同时消失,推测反应如下:
5NH4++3NO3-→4N2+9H2O+2H+
G0′=-297kJ/molNH4+
这个厌氧氨氧化(Anammox)过程的总反应是产能的,在理论上可以提供能量供微生物生长。随后Graaf又证明了这一反应是一个生物化学反应,亚硝酸盐是最合适的电子受体:
NH4++NO2-→N2+2H2O
G0′=-358kJ/molNH4+
1996年Graaf的试验研究又表明厌氧氨氧化过程是由自养菌完成的。Schmidt的试验研究则表明二氧化氮(NO2)对于厌氧氨氧化是必需的,N2O是Anammox过程的中间产物,而NO被证明对该过程有抑制作用。MarcStrous总结比较了厌氧氨氧化和好氧氨氧化的微生物生理学参数(见表3)。
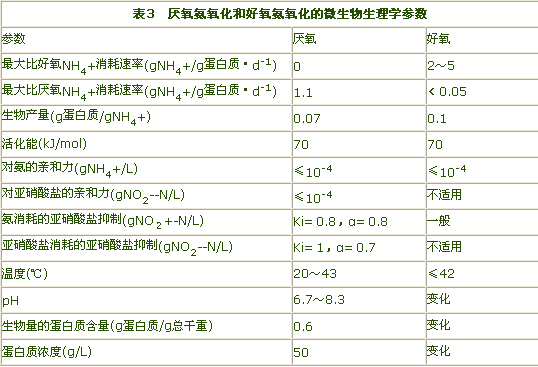 |
好氧硝化和厌氧氨氧化微生物在生理学方面是相似的,都是专属微生物,厌氧氨氧化没有好氧活动,好氧硝化微生物的厌氧活动只是厌氧氨氧化的5%。二者的生物产量、温度范围、基质亲和力、活化能大体上是相同的,只是厌氧氨氧化最大比基质转化率较低,这使得厌氧氨氧化要求较长的反应停留时间。
厌氧氨氧化在1000mg/L的氨氮或硝态氮的浓度下不会受到抑制,但是在100mg/L亚硝态氮浓度下,厌氧氨氧化过程即受到限制。亚硝酸盐抑制可以通过添加痕量厌氧氨氧化中间产物(联氨或羟氨)来克服。
目前推测厌氧氨氧化有多种途径。其中一种包括羟氨和亚硝酸盐生成N2O的反应,而N2O可以进一步转化为氮气,氨被氧化为羟氨。另一种是氨和羟氨反应生成联氨,联氨被转化成氮气并生成4个还原性[H],还原性[H]被传递到亚硝酸还原系统形成羟氨。第三种是:一方面亚硝酸被还原为NO,NO被还原为N2O,N2O再被还原成N2;另一方面,NH+4被氧化为NH2OH,NH2OH经N2H4、N2H2被转化为N2。
6 结语
总之,存在大幅度提高生物脱氮效率的生物学基础,而且效率的提高并不一定意味着成本的上升。厌氧氨氧化可以通过将好氧沉淀池出水回流到前面的厌氧反应器来完成,只是由于厌氧氨氧化的反应速度比较慢,厌氧反应器的容积可能要大一些。亚硝酸盐反硝化和好氧反硝化均要求较低的溶解氧,这样既可以节省空气量,又可以提高脱氮效率。所以在低溶解氧条件下,存在异养硝化、亚硝酸盐好氧反硝化的可能性,从而可以在节省能源和碳源的情况下实现高氨氮废水的高效脱氮,这应成为今后生物脱氮研究的重要课题。来源:中国环保频道

